The group will be welcoming a number of new PhD students later this year who have recently been awarded studentships/funding to study here. They will be:
Emily Baker, who will be joining us on an IAPETUS2 studentship. Emily will be moving from (but still working with) CEH and will be working on a project on “The validation of ecological citizen science data”, also in collaboration with the NBN and APHA. Emily will be supervised by Phil Stephens.
Eilidh Smith, who will be funded by the Whitehead Trust. Eilidh will be moving from the University of the Highlands and Islands, where she has been working on reindeer in the Cairngorms. Her focus will be a project on “Impacts of disturbance on red deer”. Eilidh will be supervised by Phil Stephens.
Peter Stewart, who will be funded by a NERC IAPETUS2 studentship. Peter will be moving from Imperial College London, to answer the question "How do invasive species impact African wild mammals?" in Kenya. Peter will be supervsied by Wayne Dawson.
Katy Ivison, who will be funded by a Leverhulme Durham ARCTIC studentship. Katy is a graduate from the University of Birmingham, and will work on a project asking "How will climate change and natural enemies affect non-native plant invasion risk in Arctic Norway?". Katy will be supervsied by Wayne Dawson.
David Jarrett is coming to Durham from the British Trust for Ornithology and is funded by a Leverhulme DurhamArctic scholarship to study for a PhD on ‘Climate change and Arctic wildlife: consequences for human-wildlife interactions & ecosystem services’.
We look forward to welcoming them to the group!
Emily Baker, who will be joining us on an IAPETUS2 studentship. Emily will be moving from (but still working with) CEH and will be working on a project on “The validation of ecological citizen science data”, also in collaboration with the NBN and APHA. Emily will be supervised by Phil Stephens.
Eilidh Smith, who will be funded by the Whitehead Trust. Eilidh will be moving from the University of the Highlands and Islands, where she has been working on reindeer in the Cairngorms. Her focus will be a project on “Impacts of disturbance on red deer”. Eilidh will be supervised by Phil Stephens.
Peter Stewart, who will be funded by a NERC IAPETUS2 studentship. Peter will be moving from Imperial College London, to answer the question "How do invasive species impact African wild mammals?" in Kenya. Peter will be supervsied by Wayne Dawson.
Katy Ivison, who will be funded by a Leverhulme Durham ARCTIC studentship. Katy is a graduate from the University of Birmingham, and will work on a project asking "How will climate change and natural enemies affect non-native plant invasion risk in Arctic Norway?". Katy will be supervsied by Wayne Dawson.
David Jarrett is coming to Durham from the British Trust for Ornithology and is funded by a Leverhulme DurhamArctic scholarship to study for a PhD on ‘Climate change and Arctic wildlife: consequences for human-wildlife interactions & ecosystem services’.
We look forward to welcoming them to the group!
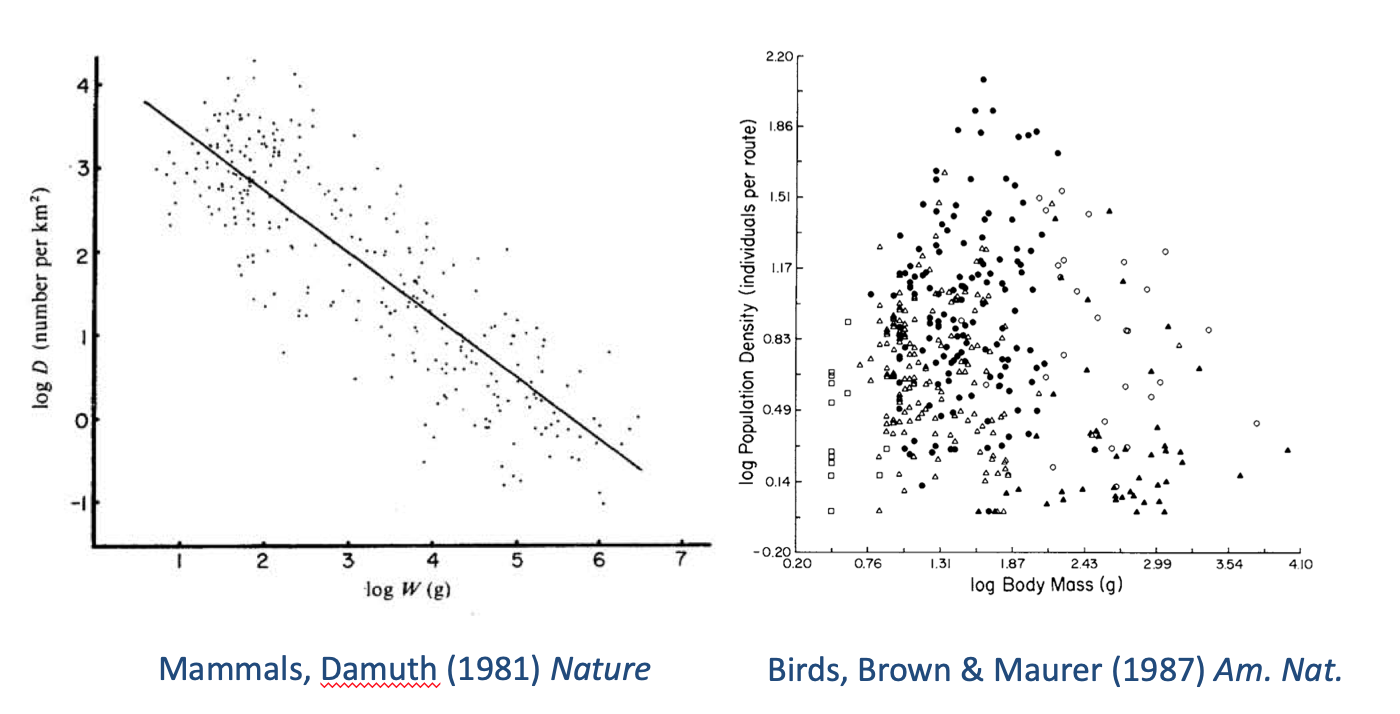
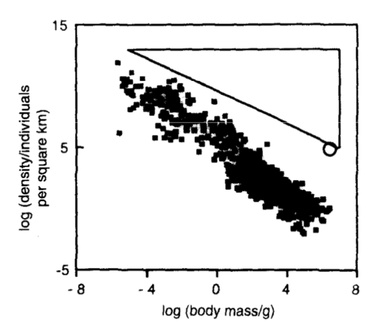
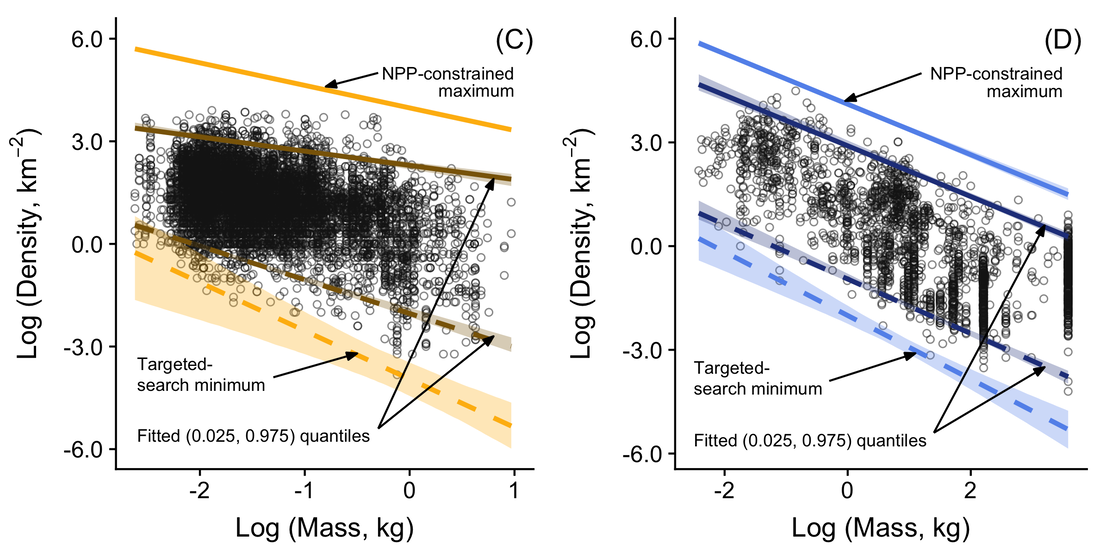
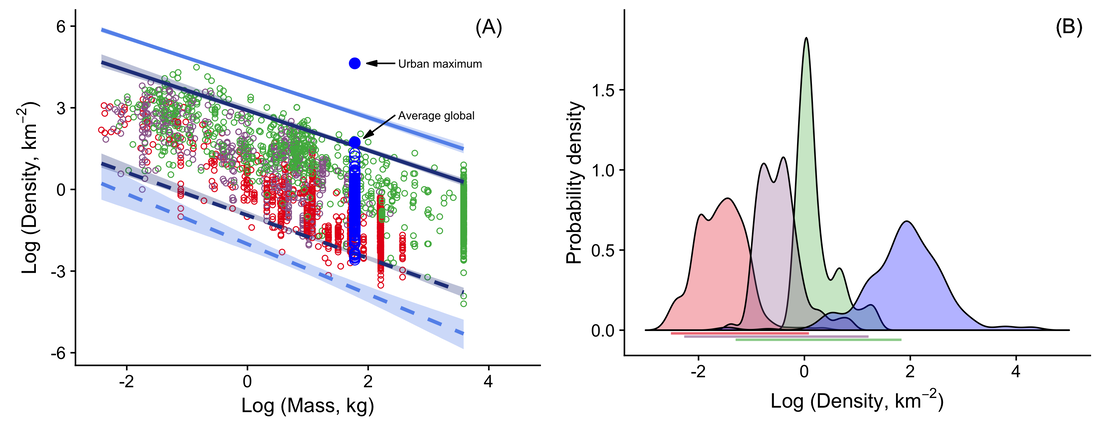
 RSS Feed
RSS Feed
